【文献解读】ceRNA机制研究经典思路
结果显示,miR-330-3p模拟物显著抑制了lnc-AIFM2-1的表达,miR-330-3p ASO增加了lnc-AIFM2-1的表达,而lnc-AIFM2-1-3‘UTR-mut抑制了它们的表达(图5D)。为研究has-miR-330-3p与CD244 mRNA之间的相互作用,将mimics NC、has-miR-330-3p mimics、ASO-NC和has-miR-330-3p ASO
ceRNA即竞争性内源RNA,是一种假说,一种全新的基因表达调控模式,即ceRNA可以通过竞争性结合miRNA的位点,从而影响miRNA与mRNA的3'UTR结合,进而影响下游mRNA的表达(ceRNA-miRNA-mRNA),影响细胞功能。这种ceRNA机制是近年来的研究热点,截至2023年5月31日,从pubmed检索ceRNA相关文章,结果有7145条,其中近三年发表的有3412篇。如此居高不下的研究热度,以及大量的高分文章,总会让人好奇关于ceRNA机制的研究是如何进行的,接下来本文将通过一篇文献来详细介绍。

题目:Lnc-AIFM2-1 promotes HBV immune escape by acting as a ceRNA for miR-330-3p to regulate CD244 expression
期刊:Frontiers in Immunology
IF:8.786
一、研究背景
慢性乙型肝炎(CHB)病毒感染是肝硬化和肝细胞癌(HCC)的主要危险因素。乙型肝炎病毒(HBV)的免疫逃逸受病毒特异性CD8+ T细胞耗竭的调控,这与负调控分子CD244的异常表达有关。然而,其潜在的机制尚不清楚。另有研究表明,非编码RNA(ncRNA),包括microRNA(miRNA)和长链非编码RNA(lncRNA),也可能在HBV感染中发挥重要的调控作用。miRNAs通过调节靶标mRNA的表达,参与了许多重要的生物学过程,如细胞信号转导和免疫反应。lncRNAs作为一类内源性竞争RNA(ceRNAs),可以作为miRNA海绵来介导基因表达。因此,本研究假设lncRNA和CD244相互作用的证据有助于阐明细胞内和细胞间分子之间的功能关系,从而为了解对HBV免疫逃逸至关重要的生物学过程、途径和相互作用网络提供见解。
二、研究思路
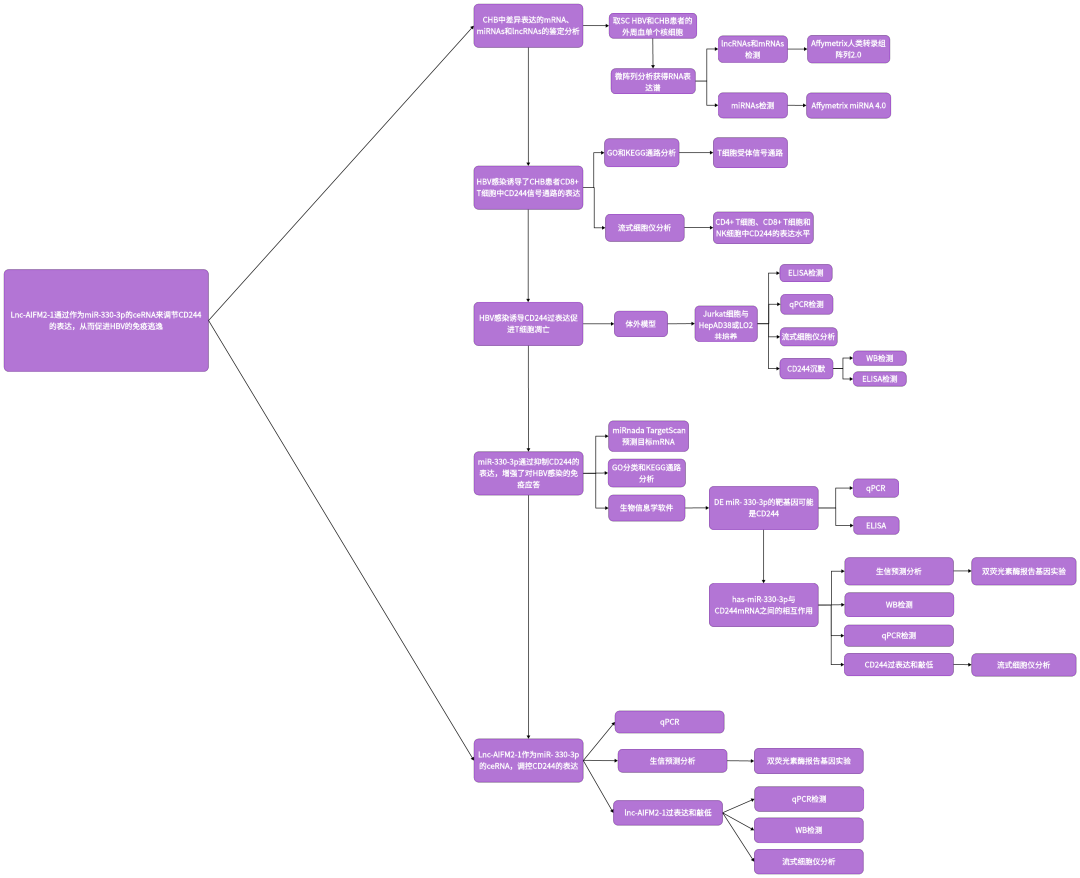
三、研究结果
1. CHB中差异表达的mRNA、miRNAs和lncRNAs的鉴定
CHB患者与SC HBV患者相比,共发现513个lncRNAs、256个mRNAs和48个miRNAs存在差异表达(DE)。其中,与SC HBV患者相比,CHB患者中368个DE lncRNAs、114个mRNAs和22个miRNAs表达上调,145个lncRNAs、142个mRNAs和26个miRNAs表达下调。火山图显示,DE RNA可以区分CHB患者和SC HBV患者(图1A-C)。
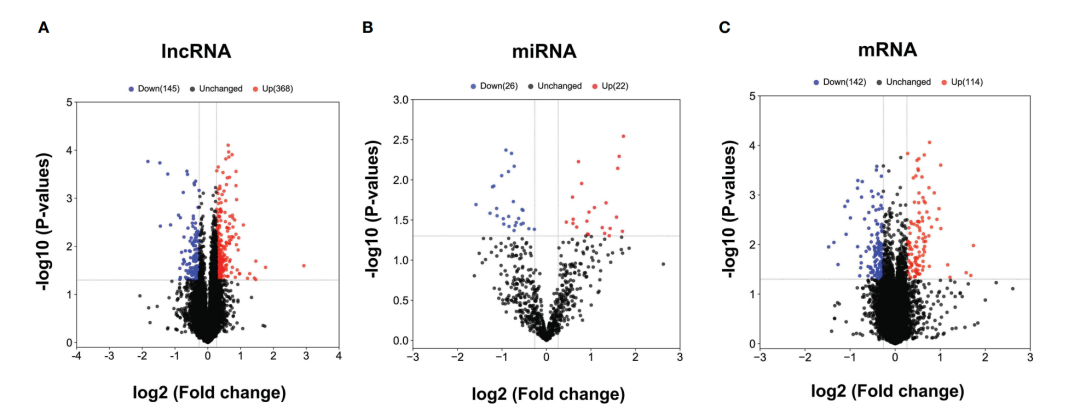
图1 CHB患者与SC HBV对照组差异表达lncRNAs、miRNAs和mRNAs的火山图。
2. HBV感染诱导了CHB患者CD8+ T细胞中CD244信号通路的表达
通过GO分析研究了DE RNA的生物学功能,这支持了GO项分析中证实的免疫应答通路的作用,包括T细胞受体信号通路(图2A-C)。为了确定CD244信号通路是否参与了抗HBV免疫应答,本研究检测了CD4+ T细胞、CD8+ T细胞和NK细胞中CD244的表达水平。流式细胞仪分析显示,与SC HBV患者相比,HBV感染诱导CD244+ CD8+ T细胞和CD244+ CD4+ T细胞显著增加,而CD244+ CD16+ NK细胞没有显著增加(图2D)。此外,无论是SC HBV患者还是CHB患者的PBMCs中,CD244+ CD8+ T细胞的百分比都远远高于CD244+ CD4+ T细胞(图2E,F)。
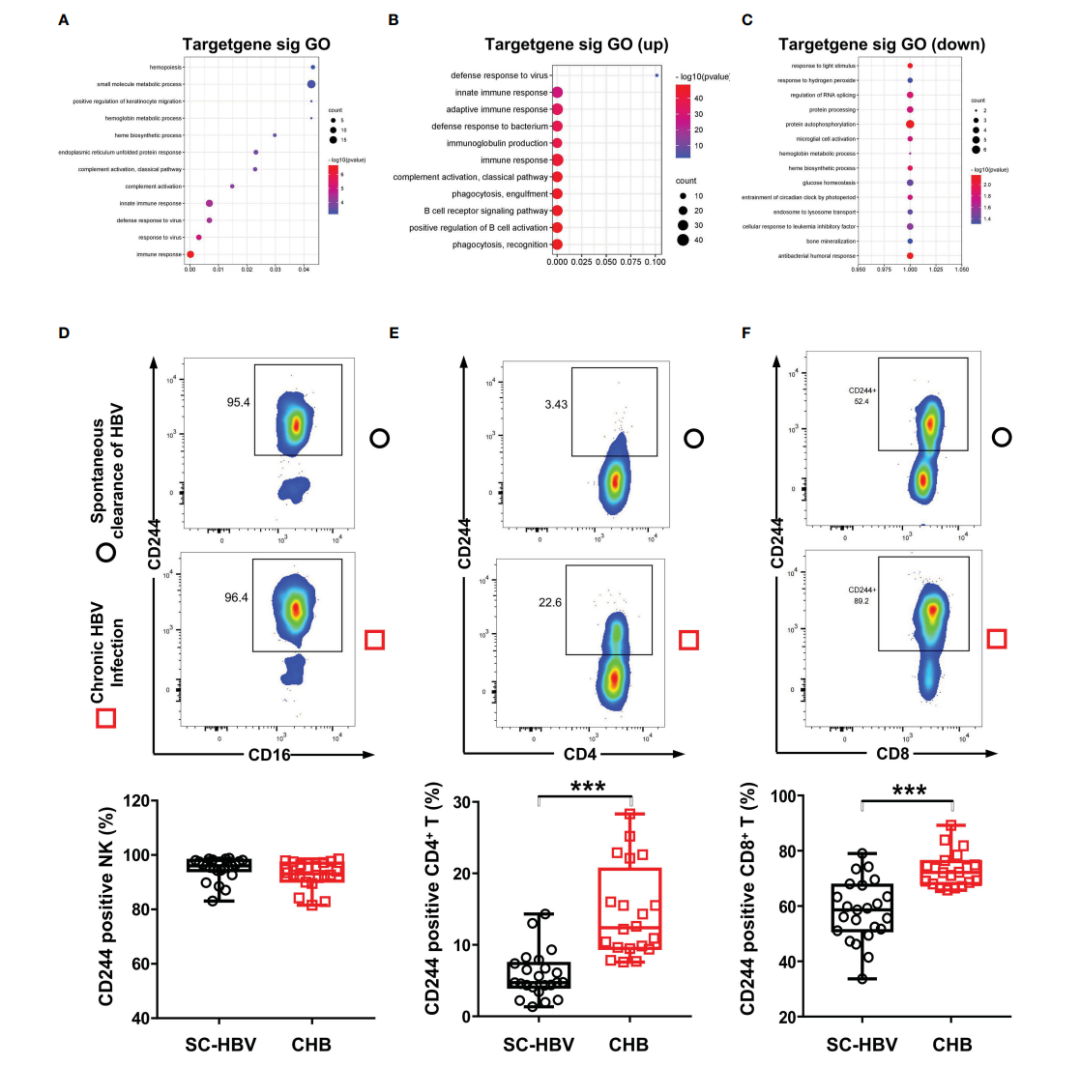
图2 CHB患者和SC HBV对照中免疫细胞上的CD244信号表达。
3. HBV感染诱导CD244过表达促进T细胞凋亡
为了明确CD244在CHB和SC HBV的T细胞中的差异表达的调控作用,本研究建立了一个体外模型。将T细胞系Jurkat细胞与正常肝细胞系LO2或感染HBV的肝细胞系HepAD38细胞共培养(图3A)。检测HBV DNA和HBsAg,以证明与Jurkat细胞共培养的HepAD38细胞中HBV的感染(图3B,C)。与Jurkat+LO2组相比,Jurkat+HepAD38组CD244 mRNA表达水平显著增加(图3D)。而与Jurkat+LO2组相比,Jurkat+HepAD38组,细胞凋亡水平增加,说明CD244的增加与T细胞凋亡有关(图3E),同时该组HBV DNA和HBsAg显著升高。相反,CD244沉默后,HBV DNA和HBsAg水平显著降低(图3F-H)。这些数据共同表明了CD244信号通路在HBV感染期间调节CD8+ T细胞免疫应答中的重要性。
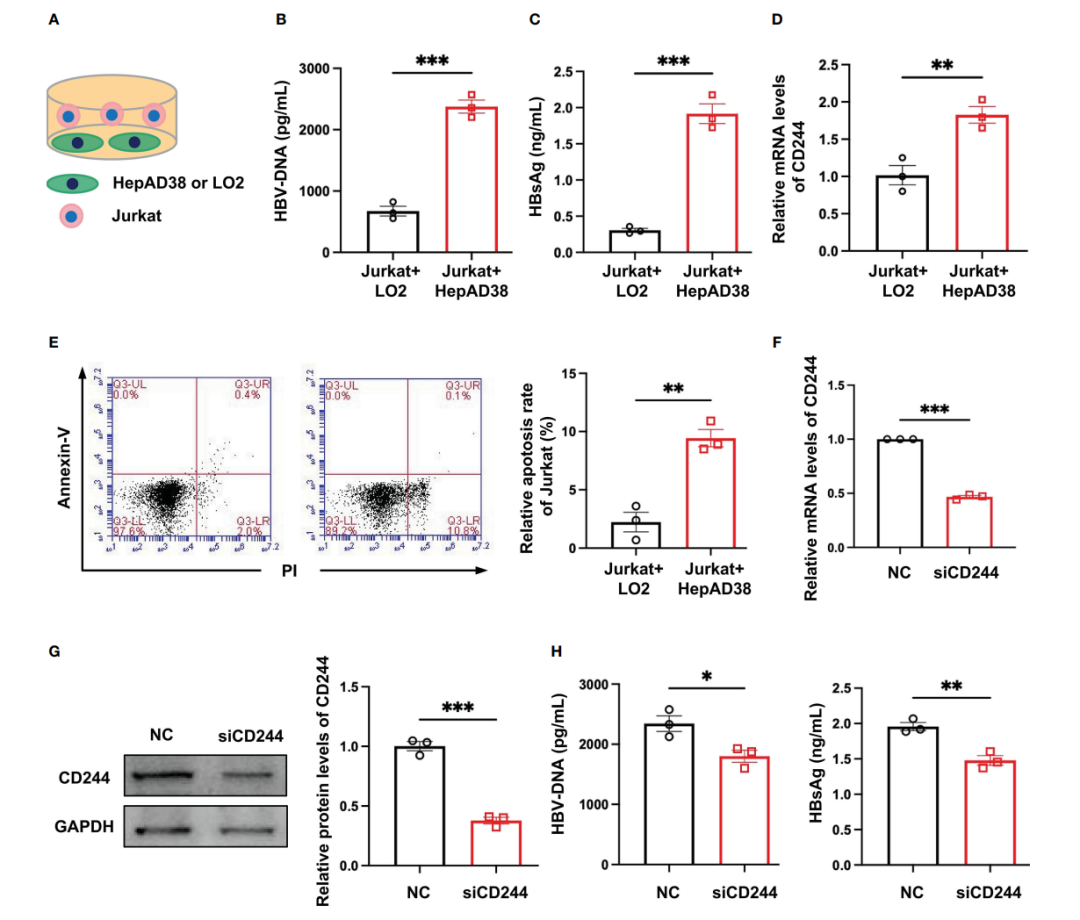
图3 CD244表达调节有或没有HBV感染的T细胞凋亡。
4. miR-330-3p通过抑制CD244的表达,增强了对HBV感染的免疫应答
为了找出调控CD244在SC HBV和CHB中差异表达的原因,本研究分析了DE miRNA。使用miRnada和TargetScan工具预测了5240个目标mRNA(图4A)。这些DE miRNAs的GO分类和KEGG通路分析显示,转录和免疫通路显著降低(图4B)。此外,生物信息学软件TargetScan预测DE miR-330-3p的靶基因可能是CD244(图4C)。然后比较CHB患者和SC HBV患者的miR-330-3p水平。RT-qPCR数据显示,CHB患者中miR-330-3p的表达明显降低(图4D)。此外,与Jurkat+LO2组相比,Jurkat+HepAD38组(HBV感染组)miR-330-3p的表达显著降低(图4E),CD244 mRNA表达水平显著升高(图3D)。相反,在与Jurkat细胞共培养的HepAD38细胞中,miR-330-3p模拟物刺激后,HBV DNA和HBsAg显著降低(图4F-H)。
为研究has-miR-330-3p与CD244 mRNA之间的相互作用,将mimics NC、has-miR-330-3p mimics、ASO-NC和has-miR-330-3p ASO分别转染与Jurkat共培养的HepAD38细胞。在证实miR-330-3p的mimics和ASO正常工作后,本研究发现miR-330-3p mimics降低了CD244的mRNA和蛋白水平,ASO miR-330-3p显著增加了CD244的表达(图4I,J)。为了进一步探讨miR-330-3p是否通过直接结合作用靶向CD244,采用双荧光素酶实验设计。结果显示,CD244-3‘UTR-wt组添加miR-330-3p mimics相比添加miR-330-3p NC萤火虫荧光素酶活性显著降低,而添加ASO miR-330-3p相比添加ASO NC萤火虫荧光素酶活性显著升高;CD244-3‘UTR-mut组无显著差异(图4K)。此外,转染CD244质粒过表达CD244,CD8+ T细胞的凋亡明显增加(图4L)。通过siRNA抑制CD244,挽救了ASO miR-330-3p诱导的T细胞的凋亡(图4L)。结果表明,has-miR-330-3p通过直接相互作用靶向CD244。
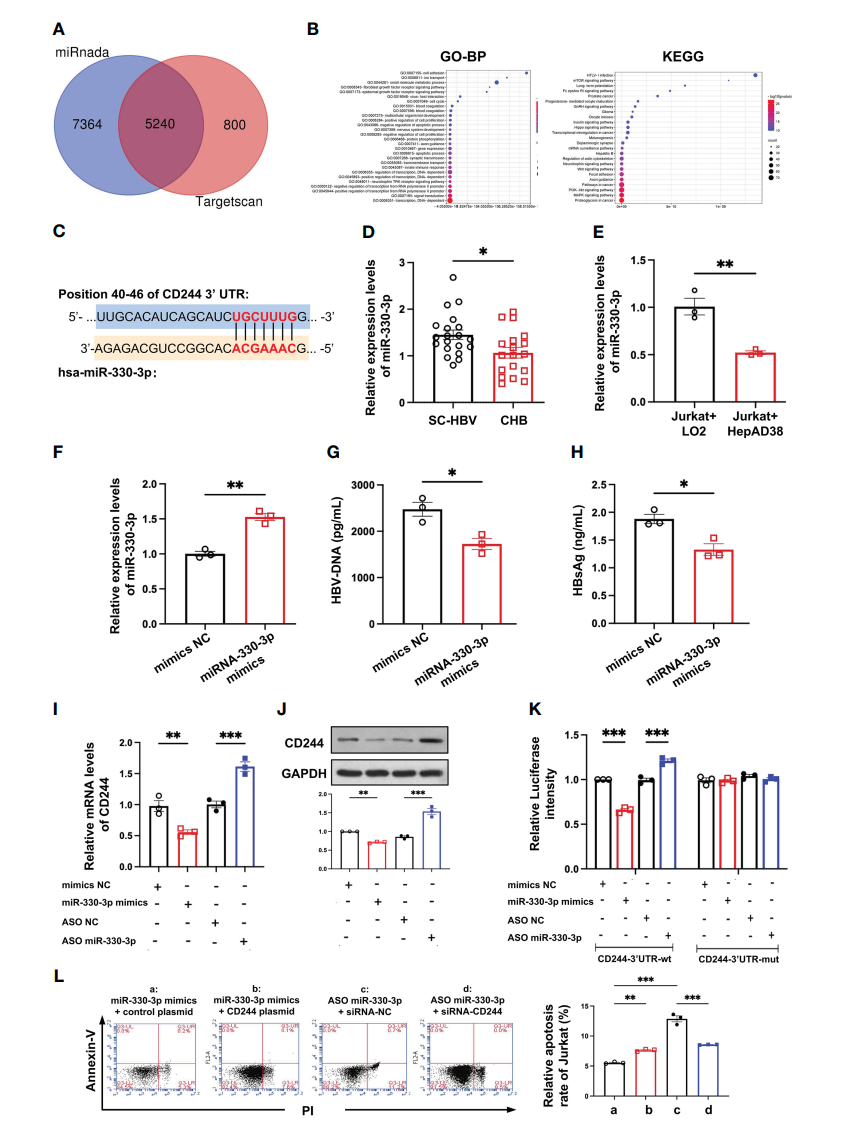
图4 miR-330-3p通过CD244改变调节对HBV感染的免疫应答。
5. Lnc-AIFM2-1作为miR- 330-3p的ceRNA,调控CD244的表达
结合lncRNA/miRNA相互作用和miRNA/mRNA相互作用,这些RNA的热图表明上调的lncRNA可以区分CHB患者和SC-HBV患者(图5A)。本研究发现lnc-AIFM2-1和CD244与miR-330-3p具有相似的结合位点(图5B)。因此,比较了CHB患者和SC HBV患者中lnc-AIFM2-1的水平。RT-qPCR数据显示,CHB患者中lnc-AIFM2-1的表达显著增加(图5C)。为了研究has-miR-330-3p和lnc-AIFM2-1之间的直接相互作用,设计了双荧光素酶实验。结果显示,miR-330-3p模拟物显著抑制了lnc-AIFM2-1的表达,miR-330-3p ASO增加了lnc-AIFM2-1的表达,而lnc-AIFM2-1-3‘UTR-mut抑制了它们的表达(图5D)。另外,通过转染lnc-AIFM2-1质粒过表达了lnc-AIFM2-1,并通过转染lnc-AIFM2-1 siRNA降低了lnc-AIFM2-1的表达(图5E)。此外,lnc-AIFM2-1的上调导致了miR-330-3p的降低和CD244的升高(图5F,G)。CD244的蛋白表达变化与mRNA一致(图5H)。此外,lnc-AIFM2-1的下调导致miR-330-3p升高,CD244水平降低(图5F,G)。过表达lnc-AIFM2-1组中CD8+ T细胞的凋亡相对过表达对照组明显增加(图5I)。相反,lncAIFM2-1 siRNA组的CD8+ T细胞凋亡相对敲低对照组减少(图5I)。此外,与lnc-AIFM2-1质粒+模拟物NC组相比,miR-330-3p模拟物的处理显著抑制了CD8+ T细胞的凋亡(图5J)。与lnc-AIFM2-1质粒+模拟物NC组相比,miR-330-3p模拟物处理后的HBV DNA和HBsAg水平也显著降低(图5K,L)。这些数据表明,lnc-AIFM2-1作为miR-330-3p的ceRNA,参与了HBV的免疫逃逸。
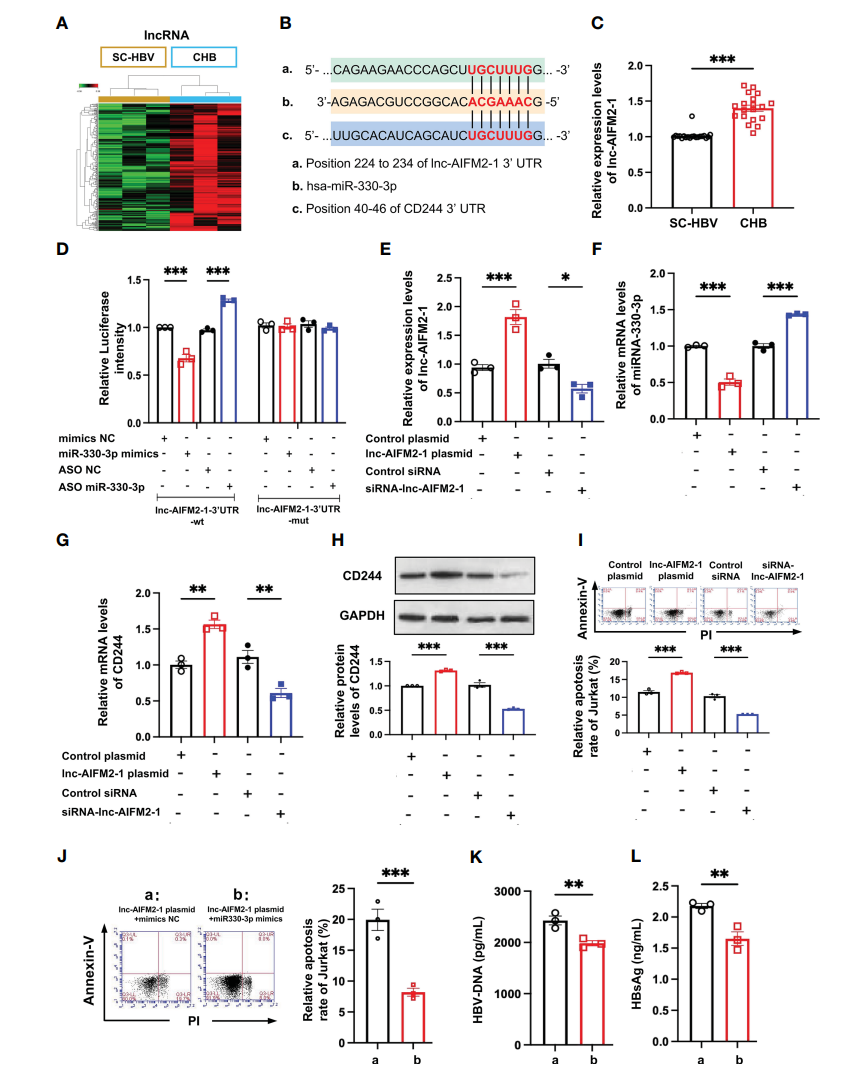
图5 lnc-AIFM2-1与miR-330-3p相互作用调节免疫应答。
原文链接:
https://www.frontiersin.org/articles/10.3389/fimmu.2023.1121795/full

有“AI”的1024 = 2048,欢迎大家加入2048 AI社区
更多推荐


 19
19 0
0- 0



 已为社区贡献5条内容
已为社区贡献5条内容

所有评论(0)